
Dimetrodon
Sobre este escolas selecção Wikipedia
SOS acredita que a educação dá uma chance melhor na vida de crianças no mundo em desenvolvimento também. Crianças SOS tem cuidado de crianças na África por 40 anos. Você pode ajudar o seu trabalho na África ?
| Dimetrodon Gama Temporal: Eopermiano , 299-270Ma | |
|---|---|
 | |
| Esqueleto montado de Dimetrodon grandis | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Família: | † sphenacodontidae |
| Subfamília: | † Sphenacodontinae |
| Género: | † Dimetrodon Cope, 1878 |
| Espécies de tipo | |
| † limbatus Dimetrodon (Cope, 1877 [limbatus originalmente clepsydrops]) | |
| Espécies | |
Veja abaixo | |
| Sinônimos | |
Bathyglyptus Case, 1911 | |
Dimetrodon ( / d aɪ m ɛ t r ə d ɒ n /; que significa "duas medidas dos dentes") é um extinto gênero de synapsid que viveu durante o Permiano Inferior , cerca de 299-270000000 anos atrás (Ma). Ele é um membro da família Sphenacodontidae. A característica mais proeminente de Dimetrodon é a grande vela em sua parte traseira formada por espinhos alongados que se estendem a partir das vértebras. Ele caminhava sobre quatro patas e tinha um crânio alto, curvado, com grandes dentes de diferentes tamanhos definidos ao longo das mandíbulas. A maioria dos fósseis foram encontrados no sudoeste dos Estados Unidos, a maioria vindo de um depósito geológico denominado Camas vermelhas no Texas e Oklahoma. Mais recentemente, fósseis foram encontrados na Alemanha. Mais de uma dúzia espécies foram nomeadas desde o gênero foi o primeiro descrito em 1878.
Dimetrodon é muitas vezes confundido como um dinossauro ou como um contemporâneo dos dinossauros na cultura popular, mas foi extinto cerca de 40 milhões de anos antes do aparecimento do primeiro dinossauro no Triássico período. Geralmente réptil-como na aparência e fisiologia, Dimetrodon é, contudo, mais estreitamente relacionado aos mamíferos do que para qualquer grupo de répteis que vivem, embora não seja um ancestral direto de qualquer mamíferos. Dimetrodon pertence a um grupo tradicionalmente chamado " répteis semelhantes a mamíferos ", mais recentemente denominado" stem-mamíferos "ou" não-mamífero synapsids "porque muitos paleontólogos de vertebrados hoje Dimetrodon grupo juntamente com mamíferos em um grupo evolutivo ou clade chamado Synapsida enquanto os dinossauros vão junto com os répteis e aves que vivem em um grupo separado, Sauropsida. Uma única abertura grande em ambos os lados da parte posterior do crânio liga Dimetrodonte com mamíferos e a distingue da maior parte dos primeiros sauropsídeos, que seja aberturas falta ou ter duas aberturas. Características tais como saliências do interior do cavidade nasal e uma crista na parte de trás da mandíbula inferior está pensado para fazer parte de uma progressão evolutiva de início tetrápodes (vertebrados de quatro ramificações) para mamíferos.
Dimetrodon era provavelmente um dos maiores predadores em ecossistemas Permiano adiantados, alimentando-se de peixes e os tetrápodes, incluindo répteis, bem como os anfíbios. As espécies menores Dimetrodon podem ter tido diferentes papéis ecológicos. A vela de Dimetrodonte pode ter sido usado para estabilizar a coluna vertebral ou para aquecer e arrefecer o corpo como uma forma de termorregulação. Alguns estudos recentes argumentam que a vela teria sido ineficaz na remoção de calor do corpo, e foi utilizada em mais provável exibição sexual.
Descrição e paleobiology
Dimetrodon era um synapsid lastreados em vela quadrúpede. A maioria das espécies Dimetrodon variaram em comprimento 1,7-3,2 metros (5,6-10 pés) e estima-se que pesava entre 28 e 250 quilogramas (62 e £ 550). A maior espécie conhecida de Dimetrodon é D. grandis em 3,2 metros (10 pés) eo menor é D. teutonis a 60 centímetros (24 polegadas). As espécies maiores de Dimetrodon estavam entre os maiores predadores do início do Permiano, embora o intimamente relacionado Tappenosaurus, conhecido a partir de fragmentos de esqueletos em rochas ligeiramente mais jovens, pode ter sido ainda maior em um número estimado de 18 pés (5,5 m) de comprimento total do corpo.
Crânio

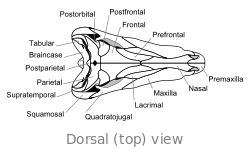
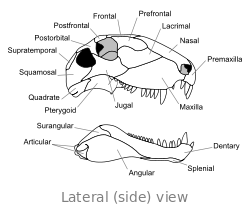
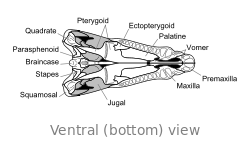
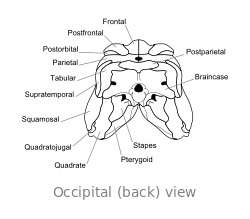
O crânio de Dimetrodon é alto e comprimido lateralmente, ou lado a lado. As órbitas estão posicionados alto e longe de volta no crânio. Por trás de cada órbita ocular é um único buraco chamado de fenestra infratemporal. Dois pares adicionais de furos pode ser visto na parte posterior do crânio quando visto de cima: a fenestra supratemporal ea fenestra posttemporal. A parte posterior do crânio (o occipital) é orientada em um pequeno ângulo para cima, uma característica que compartilha com todos os outros synapsids iniciais. A margem superior do crânio inclina para baixo em um arco convexo para a ponta da cabeça. A ponta do maxilar superior, formada pela premaxilla osso, é levantada acima da parte da maxila formada pela osso maxilar de modo a formar um "passo". maxilar Dentro deste passo é uma diastema, ou lacuna na fileira de dentes.
Dentes
O tamanho dos dentes é muito diferente ao longo do comprimento das maxilas, emprestando Dimetrodonte seu nome, o que significa "duas formas de dente" em referência a conjuntos de pequenos e grandes dentes. Um ou dois pares de caniniforms (grande pontiagudo caninos semelhantes) estendem-se desde a maxila. Grandes dentes incisivos também estão presentes nas pontas dos maxilares superiores e inferiores, enraizada na premaxillae e ossos dentário. Pequenos dentes estão presentes em torno do "passo" maxilar e atrás das caniniforms, se tornando menor mais para trás na mandíbula.
Muitos dentes são mais largos nas suas midsections e estreitar mais perto das maxilas, dando-lhes a aparência de uma lágrima. Dentes em forma de lágrima são exclusivos para dimetrodon e outros intimamente relacionados sphenacodontids, e ajudar a distingui-los de outros synapsids iniciais. Como em muitos outros synapsids início, os dentes da maioria das espécies Dimetrodon são serrilhadas em suas bordas. Os dentes de D. teutonis falta serrilhas, mas ainda tem arestas vivas.
Cavidade nasal
Na superfície interior da secção nasal de crânio são chamados nervuras nasoturbinals, que pode ter cartilagem que aumentou a área do suportados epitélio olfactivo, a camada de tecido que detecta odores. Estes cumes são muito menores do que os de synapsids posteriores a partir do final do Permiano e Triássico, cuja nasoturbinals grande são tomados como evidência para warm-bloodedness porque eles podem ter apoiado as membranas mucosas que aqueceram e umedecidos ar que entra. Assim, a cavidade nasal é de Dimetrodonte de transição entre os de vertebrados terrestres primitivas e mamíferos.
Articulação da mandíbula e do ouvido
Outra característica de transição de Dimetrodonte é uma crista na parte de trás da mandíbula chamado a lamina reflectida. A lâmina reflectida é encontrado no osso articular, que se liga ao quadrate osso do crânio para formar a articulação da mandíbula. Em antepassados de mamíferos posteriores, o articular e quadrado separado da articulação da mandíbula enquanto a articular evoluiu para a malleus osso do ouvido interno. A lâmina refletiu tornou-se parte de um anel chamado anel timpânica que suporta o tímpano em todos os mamíferos vivos.
Cauda

A cauda de Dimetrodon torna-se uma grande parte do seu comprimento total do corpo e inclui cerca de 50 vértebras caudais. Tails foram omissas ou incompletas nos esqueletos primeiro descritos de Dimetrodon; a única vértebras caudais eram conhecidos os onze mais próximo do quadril. Uma vez que estes primeiros vértebras caudais estreito rapidamente à medida que progridem mais longe do quadril, muitos paleontólogos no século XIX e início do século XX pensava que Dimetrodon tinha uma cauda muito curta. Não foi até 1927 que uma grande parte da cauda completa de Dimetrodon foi descrito.
Vela
A vela de Dimetrodon é formado por alongada espinhos neurais projectam a partir das vértebras. Cada coluna varia em forma transversal a partir de sua base para a sua ponta no que é conhecido como diferenciação "dimetrodont". Perto do corpo da vértebra, a secção transversal da coluna é comprimido lateralmente em uma forma retangular, e mais perto da ponta, ele assume uma forma em forma de oito, como uma ranhura corre ao longo de ambos os lados da coluna vertebral. A figura oito forma é pensado para reforçar a espinha, evitando flexão e fraturas. Uma secção transversal da coluna vertebral de um espécime de giganhomogenes Dimetrodon é de forma rectangular, mas conserva forma de oito anéis em forma de perto do seu centro, o que indica que a forma de espinhas podem mudar à medida que as pessoas envelhecem. A anatomia microscópica de cada coluna varia da base à ponta, indicando onde foi incorporado nos músculos das costas e onde foi exposta como parte de uma vela. Quanto menor ou porção proximal da coluna tem uma superfície rugosa que teria serviu como um ponto de ancoragem para a epaxial músculos das costas, e também tem uma rede do tecido conjuntivo denominada Fibras de Sharpey que indicam que foi incorporado dentro do corpo. Mais acima na porção distai (exterior) da coluna vertebral, a superfície do osso é mais suave. O periósteo, uma camada de tecido que envolve o osso, é coberto em pequenas ranhuras que presumivelmente suportados os vasos sanguíneos que vascularizados a vela. A grande ranhura que percorre toda a extensão da coluna vertebral já foi pensado para ser um canal para os vasos sanguíneos, mas desde que o osso não contém canais vasculares, a vela não é pensado para ter sido tão altamente vascularizado como se pensava. Alguns espécimes de Dimetrodon preservar áreas deformadas dos espinhos neurais que aparecem para ser curado-over fraturas. O osso cortical que cresceu ao longo destes intervalos é altamente vascularizado, sugerindo que o tecido macio deve ter estado presente na vela para abastecer o site com os vasos sanguíneos. Layered osso lamelar torna-se maior parte da área em corte transversal da espinha neural, e contém linhas de interrupção do crescimento que podem ser utilizados para determinar a idade de cada indivíduo no momento da morte. Em muitos espécimes as porções distais de espinhos de uma curva apertada, o que indica que a vela teria um perfil irregular na vida. Sua desonestidade sugere que tecidos moles podem não ter estendido todo o caminho até as pontas dos espinhos, o que significa que webbing da vela pode não ter sido tão extensa como é comumente imaginado.
Função de termorregulação

Os paleontólogos têm proposto muitas maneiras em que a vela poderia ter funcionado na vida. Alguns dos primeiros a pensar sobre o seu propósito sugeriu que a vela pode ter servido como camuflagem entre os juncos enquanto Dimetrodon esperou por presa, ou como uma vela real barco-like para travar o vento enquanto o animal estava na água. Outra é que os longos espinhos neurais poderia ter estabilizado o tronco, restringindo-se e para baixo movimento, o que permitiria uma circulação mais eficiente lado-a-lado durante a caminhada. Em 1940, Alfred Romer e Llewellyn Ivor Price propôs que a vela servido uma função de termorregulação, permitindo que os indivíduos para aquecer seus corpos com o calor do sol. Nos anos seguintes, muitos modelos foram criados para estimar a eficácia da termorregulação em Dimetrodon. Por exemplo, em 1973 um artigo na revista Nature, os paleontólogos CD Bramwell e PB Fellgett estimou que demorou 200 kg (440 lb) individuais sobre uma hora e meia para a sua temperatura do corpo a subir 26-32 ° C (79-90 ° F). Em 1986, Steven C. Haack concluiu que o aquecimento foi mais lento do que se pensava e que o processo provavelmente levou quatro horas. Usando um modelo baseado numa variedade de factores ambientais e aspectos fisiológicos hipótese de Dimetrodonte, Haack descobriram que a vela Dimetrodonte deixada aquecer mais rapidamente no período da manhã e atinge uma temperatura ligeiramente mais elevada do corpo durante o dia, mas que não foi eficaz em libertar o excesso de calor e não permitiu que Dimetrodon manter uma temperatura corporal mais elevado durante a noite. Em 1999, um grupo de engenheiros mecânicos criou um modelo de computador para analisar a capacidade da vela para regular a temperatura corporal durante as diferentes estações do ano, e concluiu que a vela foi benéfica para a captura e liberação de calor em todos os momentos do ano.
A maioria destes estudos dar dois papéis de termorregulação para a vela de Dimetrodonte: um como um meio de aquecimento rapidamente na parte da manhã, e o outro como uma forma de arrefecer quando a temperatura do corpo torna-se elevada Dimetrodonte e todos os outros vertebrados terrestres permianos iniciais são assumidos. ter sido a sangue-frio ou poiquilotérmicos, baseando-se o sol para manter uma temperatura corporal elevada. Por causa de seu grande tamanho, Dimetrodon teve alta inércia térmica, o que significa que mudanças na temperatura corporal ocorreu mais lentamente nele do que em animais mais pequenos-bodied. Como as temperaturas subiram no período da manhã, a presa pequeno-bodied de Dimetrodon poderia aquecer seus corpos muito mais rápido que podia algo do tamanho de Dimetrodon. Muitos paleontólogos incluindo Haack propuseram que a vela de Dimetrodon pode ter permitido que ele aqueça rapidamente na parte da manhã, a fim de manter o ritmo com sua presa. Grande área de superfície da vela também significava calor pode dissipar rapidamente para o ambiente, útil se o animal necessária para liberar o excesso de calor que foi produzido a partir de metabolismo ou que foi absorvido pelo sol. Dimetrodon pode ter angulado sua vela longe do sol para se refrescar ou o fluxo de sangue restrito à vela para manter o calor durante a noite.
Em 1986, J. Scott Turner e C. Richard Tracy propôs que a evolução de uma vela em Dimetrodon estava relacionada com a evolução do warm-bloodedness em mamíferos ancestrais. Eles pensavam que a vela de Dimetrodon lhe permitiu ser homeotérmicos, mantendo uma constante, embora, baixa temperatura corporal. Mamíferos também são homeotérmica, embora difiram em Dimetrodonte sendo endotérmico, controlando a temperatura do corpo internamente através de metabolismo aumentado. Turner e Tracy notar-se que os membros longos dos primeiros terapsídeos, um grupo mais avançado de sinapsídeos que estão intimamente relacionadas com a mamíferos, permitiu-lhes libertar o calor de uma forma semelhante à vela de Dimetrodonte. O homeothermy que se desenvolveu em animais como Dimetrodon pode ter transitado para therapsids através de uma modificação da forma do corpo, o que acabaria por tornar-se o warm-bloodedness de mamíferos.
Estudos recentes sobre a vela de Dimetrodon e outros sphenacodontids apoiar 1.986 alegação de Haack que a vela foi mal adaptado para liberar calor e manter a temperatura corporal estável. A presença de velas em espécies de pequeno-bodied de Dimetrodon, como D. milleri e D. não teutonis não caber a idéia de que o objetivo da vela foi termorregulação porque velas menores são menos capazes de transferir calor e porque pequenos corpos podem absorver e liberar o calor facilmente por conta própria. Parentes Além disso, o fecho do Dimetrodonte como Sphenacodon têm muito baixas cristas que teria sido inútil como dispositivos de termorregulação. A grande vela de Dimetrodon é pensado para ter desenvolvido gradualmente a partir de cristas menores, o que significa que a maior parte da história evolutiva da vela, termorregulação não poderia ter servido uma função importante.
A seleção sexual
Maiores espécimes bodied de Dimetrodon tem velas maiores em relação ao seu tamanho, um exemplo de alometria positiva. Alometria positiva pode beneficiar termorregulação, pois significa que, como indivíduos se tornam maiores, área de superfície aumenta mais rapidamente do que em massa. Animais maiores-corpo gerar uma grande quantidade de calor, através do metabolismo, e a quantidade de calor que tem de ser dissipado a partir da superfície do corpo é significativamente maior do que o que tem de ser dissipada pelos animais menores-corpo. Dissipação de calor eficaz pode ser previsto através de muitos animais diferentes, com uma única relação entre massa e área de superfície. No entanto, um estudo de alometria em Dimetrodon 2010 descobriu uma relação diferente entre a sua vela e massa corporal: o expoente de escala real da vela foi muito maior do que o esperado em expoente um animal adaptado à dissipação de calor. Os pesquisadores concluíram que a vela de Dimetrodon cresceu a um ritmo muito mais rápido do que o necessário para a termorregulação, e sugeriu que seleção sexual foi a principal razão para a sua evolução.
O dimorfismo sexual


Dimetrodonte pode ter sido dimorfismo sexual, o que significa que os machos e as fêmeas tiveram um pouco diferentes tamanhos de corpo. Alguns espécimes de Dimetrodon foram supor como os homens porque eles têm ossos mais espessos, velas maiores, mais longas, crânios e maxilares mais pronunciados "passos" do que outros. Com base nestas diferenças, os esqueletos montados no Museu Americano de História Natural (AMNH 4636) eo Museu Field de História Natural pode ser do sexo masculino e os esqueletos no Denver Museum of Nature and Science ( MCZ 1347) eo Universidade de Michigan Museu de História Natural pode ser fêmeas.
Espécies
| Espécies | Autoridade | Localização | Estado | Sinônimos | Imagens |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Olson, 1962 | Texas | Válido |  | |
| Dimetrodon booneorum | Romer, 1937 | Texas | Válido | ||
| Dimetrodon cruciger | Cope, 1878 | Texas | Sinônimo de Edaphosaurus cruciger | ||
| Dimetrodon dollovianus | Case, 1907 | Texas | Válido | Dollovianus Embolophorus Cope, 1888 | |
| Giganhomogenes dimetrodon | Case, 1907 | Texas | Válido |  | |
| Gigas dimetrodon | Cope, 1878 | Texas | Sinônimo de Dimetrodon grandis | Clepsydrops gigas Cope, 1878 | |
| Dimetrodon grandis | Romer e preço de 1940 | Oklahoma Texas | Válido | Clepsydrops gigas Cope, 1878 Gigas dimetrodon Cope, 1878 Theropleura grandis Case, 1907 Caso Bathyglyptus Theodori, 1911 Dimetrodon maximus Romer 1936 |  |
| Dimetrodon Incisivus | Cope, 1878 | Texas | Sinônimo de Dimetrodon limbatus | ||
| ? Dimetrodon kempae | Romer, 1937 | Texas | Possível nomen dubium | ||
| Dimetrodon limbatus | Romer e preço de 1940 | Oklahoma Texas | Válido | Clepsydrops limbatus Cope, 1877 Dimetrodon Incisivus Cope, 1878 Dimetrodon rectiformis Cope, 1878 Dimetrodon semiradicatus Cope, 1881 | |
| Dimetrodon longiramus | Case, 1907 | Texas | Sinônimo de Obtusidens secodontosaurus | ||
| Dimetrodon loomisi | Romer, 1937 | Texas Oklahoma | Válido |  | |
| Dimetrodon macrospondylus | Case, 1907 | Texas | Válido | Clepsydrops macrospondylus Cope, 1884 Dimetrodon platycentrus Case, 1907 | |
| Dimetrodon milleri | Romer, 1937 | Texas | Válido |  | |
| Dimetrodon natalis | Romer, 1936 | Texas | Válido | Clepsydrops natalis Cope, 1878 |  |
| Dimetrodon occidentalis | Berman, 1977 | Arizona Novo México Utah | Válido | ||
| Dimetrodon platycentrus | Case, 1907 | Texas | Sinônimo de Dimetrodon macrospondylus | ||
| Rectiformis dimetrodon | Cope, 1878 | Texas | Sinônimo de Dimetrodon limbatus | ||
| Dimetrodon semiradicatus | Cope, 1881 | Texas | Sinônimo de Dimetrodon limbatus | ||
| Teutonis dimetrodon | Berman et al., 2001 | Alemanha | Válido |
História Classificação
Primeiras descrições por Cope
Fósseis agora atribuídas a Dimetrodon foram estudados pela primeira vez pelo paleontólogo norte-americano Edward Drinker Cope em 1870. Cope tinha obtido os fósseis junto com aqueles de muitos outros tetrápodes Permiano (vertebrados de quatro ramificadas) de vários colecionadores que tinham vindo a explorar um grupo de rochas no Texas chamado de camas Vermelho. Entre estes coletores foram naturalista suíço Jacob Boll, Texas geólogo WF Cummins, e paleontólogo amador Charles Hazelius Sternberg. A maior parte do Cope espécimes foi para o Museu Americano de História Natural ou para o Universidade de Walker Museum de Chicago (a maior parte da coleção de fósseis Walker está agora alojado no Museu Field de História Natural).
Sternberg enviou alguns de seus próprios espécimes para paleontólogo alemão Ferdinand Broili em Universidade de Munique, embora Broili não foi tão prolífico como lidar quando se tratava de descrever espécimes. Cope rival Othniel Charles Marsh também recolheu alguns ossos de Dimetrodon, que enviou ao Museu Walker. O primeiro uso do nome Dimetrodon veio em 1878 quando Cope batizou a espécie de Dimetrodon Incisivus, rectiformis Dimetrodon, e gigas Dimetrodon na revista científica Anais da Sociedade Filosófica Americana.
No entanto, a primeira descrição de um fóssil Dimetrodon veio um ano antes, quando Cope batizou a espécie de limbatus clepsydrops das camas do vermelho do Texas. (O nome clepsydrops foi cunhado pela primeira vez por Cope em 1875 para sphenacodontid permanece de Vermilion County, Illinois, e mais tarde foi empregada para muitos espécimes sphenacontid de Texas; muitas novas espécies de sphenacodontids do Texas foram atribuídos a qualquer clepsydrops ou Dimetrodon no final do século XIX e início do século XX.) C. limbatus foi reclassificada como uma espécie de Dimetrodon em 1940, o que significa que de 1877 Cope papel foi o primeiro registro de Dimetrodon.
Cope foi o primeiro a descrever um synapsid lastreados em vela com a nomeação de clepsydrops natalis em seu artigo de 1878, embora ele chamou a vela uma barbatana e comparou-a com as cristas da moderna lagarto basilisco (Basilicus). Velas não foram preservadas nos espécimes de D. Incisivus e D. gigas que Cope descritas em seu artigo 1.878, mas espinhos alongados estavam presentes no D. rectiformis espécime ele descreveu. Cope comentou sobre a finalidade da vela em 1886, escrevendo: "A utilidade é difícil de imaginar. A menos que o animal tinha hábitos aquáticos, e nadou em suas costas, a crista ou fin deve ter sido na forma de movimentos ativos ... Os membros não são suficientemente longo, nem as garras agudas suficiente para demonstrar hábitos arbóreas, como no género existente Basilicus, onde existe uma crista semelhante. "
Descrições início do século 20
Nas primeiras décadas do século XX, paleontólogo norte-americano Caso CE autor de vários estudos sobre Dimetrodon e descreveu várias novas espécies. Ele recebeu financiamento do Carnegie Institution for seu estudo de muitos Dimetrodon espécimes em coleções do Museu Americano de História Natural e vários outros museus. Muitos destes fósseis tinha sido recolhido por Cope, mas não tinha sido completamente descrita, como era conhecido Cope para erigir novas espécies, com base em apenas alguns fragmentos ósseos.
Começando no final de 1920, o paleontólogo Alfred Romer reestudados muitos espécimes Dimetrodon e chamado várias novas espécies. Em 1940, foi co-autor Romer um grande estudo com Llewellyn Ivor Price chamado "Revisão do Pelycosauria", no qual as espécies de Dimetrodon nomeados pelo Cope e Case foram reavaliados. A maioria dos nomes de espécies consideradas válidas por Romer e preço são usadas ainda hoje.
Novas amostras
Nas décadas seguintes de Romer e preço monografia, muitos espécimes Dimetrodon foram descritas a partir de localidades fora Texas e Oklahoma. O primeiro foi descrito a partir do Four Corners região de Utah em 1966 e outro foi descrita a partir de Arizona em 1969. Em 1975, Olson relatou material de Dimetrodon de Ohio. Uma nova espécie de Dimetrodon chamado D. occidentalis (que significa "Dimetrodon ocidental") foi nomeado, em 1977, do Novo México. Os espécimes foram encontrados em Utah e Arizona, provavelmente, também pertence a D. occidentalis.
Antes dessas descobertas, uma teoria de que existia um mar agitado Midcontinental separado o que é agora Texas e Oklahoma de terras ocidentais mais durante o início do Permiano, isolando Dimetrodon a uma pequena região da América do Norte, enquanto um menor chamado sphenacodontid Sphenacodon dominaram a área ocidental. Enquanto isso seaway provavelmente existia, a descoberta de fósseis fora Texas e Oklahoma mostram que sua extensão era limitada e que não era uma barreira eficaz para a distribuição de Dimetrodon.
Em 2001, uma nova espécie de Dimetrodon chamado D. teutonis foi descrita a partir da Bacia Saar-Nahe da Alemanha, estendendo o alcance geográfico de Dimetrodon fora da América do Norte pela primeira vez.
Classificação filogenética
Dimetrodon é um dos primeiros membros de um grupo chamado synapsids, que incluem mamíferos e muitos de seus parentes extintos. É muitas vezes confundido como um dinossauro na cultura popular, apesar de ter se tornado extinto cerca de 40 milhões de anos (Ma) antes dos primeiros dinossauros apareceram. Como synapsid, Dimetrodon é mais estreitamente relacionado aos mamíferos do que é dinossauros ou qualquer réptil que vive. Nos séculos XIX e início do XX atrasados, a maioria dos paleontólogos chamado Dimetrodon um réptil de acordo com Taxonomia Linnean. No sistema de Linnean, Reptilia foi classificado como um classe e Dimetrodon foi classificado como um gênero dentro dessa classe. Apesar de uma estreita relação entre Dimetrodon e mamíferos foi apreciado na época, mamíferos pertencia a sua própria classe separada e, assim, Dimetrodon era um "mamífero-como o réptil". Os paleontólogos reconheceram que os mamíferos evoluíram a partir deste grupo em que eles chamaram uma transição réptil-to-mamífero. Mais recentemente, taxonomia filogenética tornou-se um sistema comum de classificação em paleontologia de vertebrados em que os grupos são definidos por ancestrais comuns compartilhados. Répteis e mamíferos foram colocados em subtipos, que são grupos que incluem um antepassado evolutivo comum e todos os seus descendentes. Sob sistemática filogenética, os descendentes do último ancestral comum de Dimetrodon e todos os répteis que vivem deve incluir todos os mamíferos porque Dimetrodon é agora conhecido por ser mais estreitamente relacionado aos mamíferos do que para qualquer réptil que vive. Para evitar mamíferos sendo no clado que inclui estes répteis vivos, nem Dimetrodon nem qualquer outro "mamífero-como o réptil" pode ser considerado parte desse clado. Descendentes do último ancestral comum de mamíferos e répteis (que apareceu em torno de 310 Ma na Final do Carbonífero) são, portanto, dividido em duas clades: Synapsida, que inclui Dimetrodon e mamíferos, e Sauropsida, que inclui répteis vivos e todos os répteis extintos mais estreitamente relacionados a eles do que aos mamíferos.
Dentro Synapsida, Dimetrodon é parte do clado Sphenacodontia. Sphenacodontia foi proposto pela primeira vez como um grupo synapsid início em 1940 pelos paleontólogos Alfred Romer e Llewellyn Ivor Price, juntamente com os grupos Ophiacodontia e Edaphosauria. Todos os três grupos são conhecidos do Carbonífero tardio e precoce Permian. Romer e Preço os distinguia principalmente por postcranial características, tais como as formas de membros e vértebras. Ophiacodontia foi considerado o grupo mais primitivo porque os seus membros apareceram mais reptiliano, e Sphenacodontia era o mais avançado porque seus membros apareceram mais como um grupo chamado Therapsida, que incluiu os parentes mais próximos de mamíferos. Romer e Preço colocado um outro grupo de primeiros syapsids chamado varanopids dentro Sphenacodontia, considerando-os mais primitivos do que outros sphenacodonts como Dimetrodon. Eles pensaram varanopids e Dimetrodon -como sphenacodonts estavam intimamente relacionados, pois ambos os grupos eram carnívoros, embora varanopids são muito menores e mais lagarto-like, faltando velas. A visão moderna das relações sinapsídeos foi proposto pelo paleontólogo Robert R. Reisz em 1986, cujo estudo incluía recursos encontrados principalmente no crânio e não no esqueleto pós-craniano. Dimetrodon ainda é considerado um sphenacodont ao abrigo do presente filogenia, mas varanodontids são agora considerados mais synapsids basais que não se enquadrem Sphenacodontia. Dentro Sphenacodontia é o grupo Sphenacodontoidea, que por sua vez contém Sphenacodontidae e Therapsida. Sphenacodontidae é o grupo que contém Dimetrodon e vários outros synapsids lastreados em vela como Sphenacodon e Secodontosaurus, enquanto Therapsida inclui mamíferos e seus principalmente Permiano e Triássico parentes. Abaixo está uma cladogram modificado a partir da análise de Benson (no prelo), que segue esta filogenia:
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecologia

Os fósseis de Dimetrodon são conhecidos dos Estados Unidos (Texas, Oklahoma, Novo México, Arizona, Utah e Ohio) e na Alemanha, áreas que eram parte do supercontinente Euramerica durante o Permiano Inferior. Nos Estados Unidos, quase todo o material atribuído a Dimetrodon veio de três grupos geológicos no Texas centro-norte e centro-sul de Oklahoma: o Limpar Grupo Fork, o Wichita Group, ea Pearce Grupo River. A maioria dos achados fósseis fazem parte de ecossistemas de várzea que, durante o Permiano, teria sido vastas zonas húmidas. Em particular, o camas de Texas Red é uma área de grande diversidade de fósseis tetrápodes, ou vertebrados de quatro ramificações. Além de Dimetrodon, os tetrápodes mais comuns nas camas do vermelho e em todo depósitos do Permiano adiantados no sudoeste dos Estados Unidos são os anfíbios Archeria, Diplocaulus, Eryops, e Trimerorhachis, o reptiliomorph Seymouria, o réptil Captorhinus, e os synapsids Ophiacodon e Edaphosaurus. Estes tetrápodes constituído um grupo de animais que paleontólogo Everett C. Olson chamado de "chronofauna Permo-Carbonífero," um fauna que dominaram o ecossistema Euramerican continental há vários milhões de anos. Com base na geologia de depósitos como as camas do vermelho, a fauna é pensado para ter habitado a planície bem-vegetadas deltaic ecossistema.
Comida web
Olson fez muitas inferências sobre a paleoecologia das camas Texas vermelhos eo papel da Dimetrodon dentro de seu ecossistema. Ele propôs várias principais tipos de ecossistemas em que os primeiros tetrápodes viviam. Dimetrodon pertencia ao ecossistema mais primitiva, que se desenvolveu a partir de cadeias alimentares aquáticas. Nele, plantas aquáticas foram o primária produtores e foram em grande parte alimentada por cima do peixe e invertebrados aquáticos. A maioria dos vertebrados terrestres alimentados com esses consumidores primários aquáticos. Dimetrodon era provavelmente a predador de topo do ecossistema camas do vermelho, alimentando-se de uma variedade de organismos, como a grande peixe Xenacanthus, os anfíbios aquáticos e Trimerorhachis diplocaulus, e os tetrápodes terrestres e Seymouria Trematops. Insetos são conhecidos a partir das camas Eopermiano vermelhas e provavelmente foram envolvidos em algum grau na mesma teia alimentar como Dimetrodon, alimentando pequenos répteis como Captorhinus. A assembléia Camas vermelhas também incluiu alguns dos primeiros grandes herbívoros que vivem na terra como edaphosaurus e Diadectes. Alimentar principalmente em plantas terrestres, esses herbívoros não derivam sua energia a partir de cadeias alimentares aquáticas. De acordo com Olson, o melhor análogo moderno para o Dimetrodon ecossistema habitado é o Everglades.
As únicas espécies de Dimetrodon foram encontrados fora do sudoeste dos Estados Unidos é Dimetrodon teutonis da Alemanha. Os seus restos foram encontrados no Formação Tambach em um site fóssil chamado a localização Bromacker. Assemblage de tetrápodes Permiano adiantados do Bromacker é incomum em que há poucas synapsids de grande porte que servem o papel de predadores de topo. D. teutonis é estimada para ter sido de apenas 1,7 metros (5,6 pés) de comprimento, muito pequeno para presa na grande herbívoros diadectid que são abundantes na assembléia Bromacker. É mais provável que comeram pequenos vertebrados e insetos. Apenas três fósseis pode ser atribuída a grandes predadores, e eles são pensados para ter sido quer grandes ou pequenas varanopids sphenacodonts, os quais poderiam potencialmente presas em D. teutonis. Em contraste com as camas do vermelho deltáicos de várzea do Texas, os depósitos Bromacker são pensados para ter representado um ambiente de sequeiro, sem espécies aquáticas. É possível que os carnívoros de grande porte não faziam parte do assemblage Bromacker porque eram dependentes de grandes anfíbios aquáticos para a alimentação.
Os juvenis
Embora algumas espécies Dimetrodon poderia crescer muito grande, muitos espécimes juvenis são conhecidos. Paleontólogo Robert T. Bakker sugeriu em 1982 que adultos Dimetrodon habitada várzeas abertas enquanto que os juvenis preferido swamplands e margens do lago mais isolado. Esses ambientes foram inferidas a partir do tipo de sedimento em que foram encontrados ossos Dimetrodon. Bakker pensou que os jovens Dimetrodon teria se comportado de forma semelhante aos juvenis de viver grandes répteis em evitar os adultos de sua própria espécie. Em répteis que vivem, os adultos podem competir pelos mesmos recursos como seus jovens e pode até rapina sobre eles. No entanto, um estudo posterior por Donald Brinkman lançar dúvidas sobre a idéia de Bakker, mostrando que os espécimes encontrados em sedimentos de várzea e pântano representado duas espécies diferentes, não as populações adulta e juvenil da mesma espécie.