
Massospondylus
Informações de fundo
Crianças SOS tentou tornar o conteúdo mais acessível Wikipedia por esta selecção escolas. Você quer saber sobre o patrocínio? Veja www.sponsorachild.org.uk
| Massospondylus Intervalo temporal: Jurássico Inferior, 200-183Ma | |
|---|---|
 | |
| Impressão do artista de Massospondylus descreve o animal como bípede | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Classe: | Reptilia |
| Ordem: | Saurischia |
| Clado: | † Sauropodomorpha |
| Clado: | † plateosauria |
| Clado: | † massopoda |
| Família: | † massospondylidae |
| Género: | † Massospondylus Owen, 1854 |
| Espécies | |
| Sinônimos | |
| |
Massospondylus (pron .: / ˌ m æ s ɵ s p ɒ n d ɨ l ə s / mas-oh- SPON di-ləs; a partir de grego , μάσσων (Masson, "mais") e σπόνδυλος (spondylos, "vértebra")) é um género de prosauropod dinossauro do início do jurássico Período ( Hettangian para Pliensbaquiano idades, ca. 200-183 milhões de anos atrás). Ele foi descrito por Sir Richard Owen, em 1854, a partir de restos encontrados em África do Sul , e é, portanto, um dos primeiros dinossauros ter sido nomeado. Os fósseis foram encontrados desde em outros locais na África do Sul, Lesotho e Zimbabwe . O material do Arizona de Formação Kayenta, Índia e Argentina foi atribuído a este gênero em vários momentos, mas o Arizonan e materiais argentino estão agora atribuídos a outros gêneros.
O Tipo de espécies é M. carinatus; outras sete espécies foram nomeados durante os últimos 150 anos, mas somente M. kaalae entre estes ainda é considerado válido. Prossaurópode sistemática foram objecto de diversas revisões durante os últimos anos, e muitos cientistas discordam onde exatamente Massospondylus encontra-se na árvore evolutiva dos dinossauros. O nome da família Massospondylidae uma vez foi cunhado para o gênero, mas porque o conhecimento das relações prossaurópode está em um estado de fluxo, não é claro que outros dinossauros, se existem alguns pertencem a um agrupamento natural de massospondylids; vários trabalhos 2,007 apoiar a validade da família.
Embora Massospondylus foi descrito como longo quadrúpede, um estudo de 2007 descobriu que ele seja bípede. Provavelmente era um comedor de plantas ( herbívoros ), embora especula-se que os prossaurópodes pode ter sido onívora. Este animal, 4-6 metros (13-20 pés) de comprimento, tinha um pescoço comprido e cauda, com uma cabeça pequena e corpo esbelto. Em cada uma de suas patas dianteiras, ela deu à luz um polegar garra afiada que foi usado na defesa ou alimentação. Estudos recentes indicam Massospondylus cresceu de forma constante ao longo de sua vida útil, possuído sacos de ar semelhantes aos dos pássaros, e pode ter se importado para a sua jovem.
Descrição
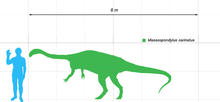
Massospondylus era uma de médio porte prosauropod que era de cerca de 4 metros (13 pés) de comprimento e pesava cerca de 135 kg (300 libras), embora algumas fontes estimam seu comprimento de até 6 metros (20 ft). Foi um prosauropod típico com um corpo esbelto, um longo pescoço e uma proporção muito pequena cabeça. A coluna vertebral foi composto por nove cervical (pescoço) vértebras, 13 dorsal (costas) vértebras, três sacral (quadril) vértebras, e, pelo menos, 40 caudal (cauda) vértebras. O pubis olhou para a frente, como acontece com a maioria dos saurischians . Ele tinha uma compilação mais leve que a de Plateosaurus , um dinossauro prosauropod outra forma similar. O pescoço foi proporcionalmente maior do que na maioria dos outros prossaurópodes, com as cervicais lugar, sendo quatro vezes mais tão grande. Os membros anteriores foram apenas metade do comprimento dos membros posteriores, mas bastante poderosa, tal como indicado pela ampla extremidade superior do úmero que fornecida áreas de fixação para um grande braço musculatura. Como Plateosaurus, que tinha cinco dedos em cada mão e pé. A mão foi curto e largo, com uma grande foice polegar em forma de garra utilizado para a alimentação ou para defesa contra predadores. O polegar foi o dedo mais longo na mão, enquanto o quarto e quinto dígitos foram pequenas, dando as patas dianteiras de um olhar torto.
Anatomia craniana
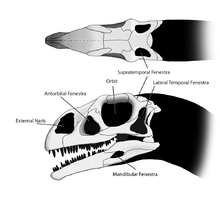
A pequena cabeça de Massospondylus foi de aproximadamente metade do comprimento do fêmur. Numerosos aberturas, ou fenestrae, no crânio reduziu o seu peso e desde que haja espaço para a fixação do músculo e dos órgãos sensoriais. Estas fenestrações estavam presentes em pares, um em cada lado do crânio. Na parte da frente do crânio eram dois grandes, elíptica narinas, que eram aproximadamente metade do tamanho das órbitas. As órbitas eram proporcionalmente maior em Massospondylus que em géneros relacionados como o Plateosaurus. O antorbital fenestrae, menor do que as observadas em Plateosaurus, situavam-se entre os olhos e do nariz. Na parte traseira do crânio eram mais dois pares de fenestrae temporais: o fenestrae temporal, lateral, imediatamente atrás das órbitas oculares, que tinham a forma de um "T" invertido em Massospondylus, e o fenestrae supratemporal no topo do crânio. Pequeno fenestrae também penetrou cada mandíbula. A forma do crânio é tradicionalmente restaurado como mais larga e mais curta do que a de Plateosaurus, mas este aspecto pode ser devido apenas ao diferencial esmagamento experimentada pelos vários espécimes. Algumas características do crânio são variáveis entre os indivíduos; por exemplo, a espessura da borda superior da órbita e a altura do posterior maxila. Estas diferenças podem ser devido a dimorfismo sexual ou variação individual.
Contagem de dente é variável entre os indivíduos e aumenta com o tamanho do crânio. O premaxilla mostra um número constante de 4 dentes por lado em todos os crânios conhecidos, mas na maxila intervalos de contagem de dentes a partir de 14 a 22. Existem 26 dentes em cada um dos lados da mandíbula inferior do crânio conhecido o maior. A altura dos dentes diminui coroas de frente para trás na maxila superior, mas foi mais ou menos constante na mandíbula inferior. A falta de desgaste do dente pronunciada e a altura variável das coroas sugere que os dentes foram substituídos por novos sucedendo em intervalos de tempo relativamente curtos. Notavelmente, houve variação da morfologia dos dentes com base na posição dos dentes no maxilar. O heterodonty presente em Massospondylus é maior do que a presente no Plateosaurus , embora sem surpresa não tão pronunciada como a especialização dos dentes Heterodontosaurus. Os dentes mais perto da frente do focinho tinha secções transversais redondas e afunilada para pontos, ao contrário dos dentes posteriores, que eram espatulado e teve secções transversais ovais.
Tal como acontece com outros prossaurópodes, tem sido proposto que Massospondylus tinha bochechas. Esta teoria foi proposta porque existem alguns grandes buracos para os vasos sanguíneos nas superfícies dos ossos da mandíbula, ao contrário dos inúmeros buracos pequenos presentes nas mandíbulas dos répteis cheekless. As bochechas teria impedido comida de derramar fora quando Massospondylus comeu. Crompton e Attridge (1986) descrito crânios de Massospondylus como possuindo pronunciada overbites e sugeriu a presença de um bico córneo na ponta da mandíbula inferior para compensar a diferença de comprimento e são responsáveis por desgaste dental sobre os dentes na ponta do focinho. No entanto, a diferença de comprimento pode ser uma má interpretação com base em esmagamento em um avião de cima para baixo, ea posse de um bico é considerada improvável em estudos recentes.
Descoberta
Os primeiros fósseis de Massospondylus foram descritos por paleontólogo Sir Richard Owen em 1854. Originalmente, Owen não reconhecer esses achados como as de um dinossauro; em vez disso ele atribuiu-os a "grandes, extinto, répteis carnívoros" que foram relacionados a de hoje lagartos, camaleões e iguanas. Este material, uma coleção de 56 ossos, foi encontrado em 1853 pelo agrimensor governo Joseph Millard Orpen no Formação superior em Elliot Harrismith, África do Sul e foi doado ao Museu Hunterian no Royal College of Surgeons em Londres. Entre os restos eram vértebras do pescoço, costas e cauda; uma lâmina de ombro; um úmero; uma pélvis parciais; um fêmur; um tíbia; e ossos das mãos e dos pés. Todos estes ossos foram encontrados desarticulado, o que torna difícil determinar se todo o material pertence a uma única espécie ou não. No entanto, Owen foi capaz de distinguir três tipos diferentes de vértebras caudais, que se atribui a três géneros diferentes:. Pachyspondylus, Leptospondylus e Massospondylus Massospondylus foi separado dos outros dois géneros com base na sua vértebras caudais muito mais tempo, o que também conduzem a o nome científico que tem sido derivada dos gregos termos Masson / μάσσων 'mais' e spondylos / σπόνδυλος 'vértebra'. No entanto, mais tarde verificou-se que o putativo vértebras caudais de Massospondylus foram realmente vértebras cervicais e que todo o material pertence provavelmente apenas para uma espécie chamusque. Em 10 de maio de 1941, o Museu Hunterian foi demolida por uma bomba alemã, destruindo todos os fósseis; unicamente elencos permanecem.
Possíveis Massospondylus restos foram encontrados na formação superior de Elliot, a Formação clarens, eo Bushveld Sandstone da África do Sul e Lesoto ; o Floresta Arenito ea Alta Karroo Sandstone de Zimbabwe ; e o Formação de Kayenta Arizona. Estes restos consistem em pelo menos 80 esqueletos parciais e quatro crânios, representando ambos os jovens e adultos. O relatório de Massospondylus de Formação Kayenta do Arizona é baseado em um crânio descrito em 1985. O crânio do espécime Kayenta do Arizona é 25% maior do que o maior crânio de qualquer espécime Africano. O espécime Kayenta possui quatro dentes na premaxilla e dezesseis no maxila. Excepcionalmente entre dinossauros, também teve pequena, one-milimétrica (0,04 in-) palatal dentes longa. Reestudo recente de crânios Massospondylus Africano, no entanto, indica que a amostra Kayenta não pertence ao Massospondylus. Este crânio Kayenta e elementos postcranial associados, identificadas em conjunto pela MCZ 8893, foi indicado ao gênero recém-descrita Sarahsaurus.
Massospondylus também haviam sido notificados da Argentina , mas esta foi reavaliada como um gênero intimamente relacionado, mas distinto. Os fósseis incluiu diversos esqueletos parciais e pelo menos um crânio, encontrado no Jurássico Inferior Formação Canon del Colorado de San Juan, Argentina. Este material foi nomeado Adeopapposaurus em 2009.
Espécies

Muitas espécies foram nomeados, embora a maioria não são mais considerados válidos. M. carinatus, nomeado por Richard Owen, é a espécies de tipo. Outras espécies nomeadas incluem: M. browni ( Seeley, 1895), M. harriesi ( Broom 1911), M. hislopi ( Lydekker, 1890), M. huenei (Cooper, 1981), M. kaalae (Barrett 2009), M. rawesi (Lydekker, 1890), e M. schwarzi ( Haughton, 1924).
M. browni, M. harriesi, e M. schwarzi foram todos encontrados na Formação Superior de Elliot Província do Cabo, África do Sul. Todos eles se baseiam em material fragmentado, e foram considerados indeterminado na revisão mais recente. M. browni baseia-se em dois cervical, dois para trás, e três caudal vértebras e diversos elementos dos membros posteriores. M. harriesi é conhecido a partir de um membro anterior bem preservados e partes de um dos membros posteriores. M. schwarzi é conhecido a partir de um membro posterior e incompleta sacro. M. hislopi e M. rawesi foram nomeados a partir de fósseis encontrados na Índia . M. hislopi baseia-se nas vértebras da Triássico Superior Formação de Måleri Andhra Pradesh, enquanto M. rawesi baseia-se num dente do Formação Cretáceo Superior de Takli Maharashtra. M. hislopi foi retido tentativamente como um sauropodomorph indeterminado na última revisão, mas M. rawesi pode ser um terópode ou nondinosaur. M. huenei é uma combinação derivada por Cooper para Lufengosaurus huenei, como ele considerava Lufengosaurus e Massospondylus ser sinônimos. Este sinonímia não é mais aceita.
M. kaalae foi descrita em 2009 com base em um crânio parcial da Formação Superior em Elliot Eastern Cape da África do Sul. Esta espécie é conhecido a partir do mesmo tempo e na região de algumas amostras de M. carinatus. Ela difere das espécies de tipo na morfologia da caixa craniana, bem como em vários outros caracteres do crânio, como as proporções de a pré-maxila.
Nomes duvidosos
Vários dinossauros são muitas vezes consideradas sinônimo de Massospondylus. Estes incluem o Leptospondylus acima indicado e Pachyspondylus bem como Aristosaurus, Dromicosaurus, Gryponyx taylori e Hortalotarsus, que são nomes duvidosos de pouco valor científico. Hortalotarsus skirtopodus foi nomeado por Harry Seeley em 1894. De acordo com Broom (1911), "Originalmente a maior parte do esqueleto estava no rock, e foi considerado pelos agricultores como o esqueleto de um Bushman, mas diz-se ter sido destruído por medo de que um Bushman Esqueleto na rocha pode tendem a enfraquecer a crença religiosa da nova geração. " Alguns ossos da perna parciais foram resgatados. Juntamente com Massospondylus carinatus, Owen nomeado Leptospondylus capensis e Pachyspondylus orpenii. Aristosaurus erectus foi nomeado por ECN van Hoepen em 1920 com base em um esqueleto quase completo. Hoepen também chamado Dromicosaurus gracilis, que consistiu de um esqueleto parcial. Gryponyx taylori foi nomeado por Sidney H. Haughton em 1924. É composto por ossos do quadril. Todos os fósseis acima vêm dos estágios de fauna Hetangiano ou Sinemurian da África do Sul, onde Massospondylus foi encontrado. Segundo as regras de nomenclatura zoológica, esses nomes são sinônimos juniores. Eles foram nomeados após Massospondylus foi descrita em um artigo científico; o nome Massospondylus leva, assim, prioridade.
Ignavusaurus, conhecido a partir de um espécime jovem, também pode ser sinônimo de Massospondylus.
Classificação
|
Massospondylus é um prosauropod, um agrupamento de início saurischian dinossauros que viveram durante o Triássico e Jurássico, mas que tinha morrido, até ao final do Jurássico. Outros membros do grupo incluem Plateosaurus , Yunnanosaurus, e Riojassauro. Sistemática sauropodomorph basais continuam a sofrer de revisão, e uma vez que muitos géneros considerados prossaurópodes clássicos recentemente foram removidos a partir do grupo em nomenclatura filogenética, com o fundamento de que a sua inclusão não constitui um clado (um agrupamento natural que contém todos os descendentes de um único ancestral comum). Exatamente que os animais constituem uma monophyletic (agrupamento natural) de prossaurópodes é clara. Yates e Kitching (2003) publicou um clado consistindo de riojassauro, Plateosaurus, Coloradisaurus, Massospondylus, e Lufengosaurus. Galton e Upchurch (2004) incluído ammosaurus , Anchisaurus, Azendohsaurus, Camelotia, coloradisaurus, Euskelosaurus, Jingshanosaurus, Lessemsaurus, Lufengosaurus, Massospondylus, Melanorossauro, Mussauro, Plateosaurus, riojassauro, Ruehleia, Saturnalia, Sellosaurus , Thecodontosaurus, Yimenosaurus e Yunnanosaurus em um Prosauropoda monofilético. Wilson (2005) considerou Massospondylus, jingshanosaurus, Plateosaurus, e Lufengosaurus um grupo natural, com Blikanasaurus e Antetonitrus possíveis saurópodes. Bonnan e Yates (2007) considerou Camelotia, Blikanasaurus e melanorossauro possíveis saurópodes. Yates (2007) colocou antetonitrus, melanorossauro, e Blikanasaurus como saurópodes basais e se recusou a usar o termo Prosauropoda, como ele considerou sinônimo de Plateosauridae. No entanto, ele não descartou a possibilidade de que um pequeno grupo de prossaurópodes consistindo de Plateosaurus, riojassauro, Massospondylus e seus parentes mais próximos eram monofilético.
Massospondylus é a Tipo de Gênero da família proposta Massospondylidae, a que dá o seu nome. A família massospondylidae pode também incluir Yunnanosaurus, embora Lu et al (2007). Yunnanosaurus colocados na sua própria família. Yates (2007) considerou Massospondylus, coloradisaurus, e massospondylids Lufengosaurus, com Yunnanosaurus em Anchisauria. Smith e Pol (2007) também encontraram uma massospondylidae em sua análise filogenética, incluindo Massospondylus, coloradisaurus, e Lufengosaurus, bem como o seu novo gênero, Glacialisaurus. Adeopapposaurus, com base nos fósseis que se pensava pertencer a uma forma Sul-Americana de Massospondylus, também foi classificado como um massospondylid, como foi Leyesaurus, outro gênero sul-americano que foi nomeado em 2011. Pradhania foi originalmente considerada como um basal sauropodomorph mas a nova análise cladística realizada por Novas et al., 2011 sugere que é um Pradhania massospondylid. Pradhania apresenta dois traços comuns da massospondylidae recuperado em sua análise filogenética, e os fósseis de Pradhania foram descobertos a partir da mesma região e bacia na Índia como M. hislopi.
Paleoecologia
O faunas e floras do jurássico adiantado foram semelhantes em todo o mundo, com coníferas adaptados para o tempo quente se tornando as plantas comuns, e prossaurópodes e basais terópodes os principais constituintes de uma fauna de dinossauros no mundo inteiro. O ambiente do início de Jurassic África Austral tem sido descrito como um deserto. Massospondylus africano foi contemporâneo de temnospondyli; tartarugas ; um Sphenodontia; rauisuchia; cedo crocodilomorfos; tritylodontid e trithelodontid therapsids; morganucodontid mamíferos ; e dinossauros, incluindo o pequeno terópode Megapnosaurus rhodesiensis e vários gêneros de primeiras ornithischians , como Lesothosaurus eo heterodontosaurids Abrictosaurus, Heterodontosaurus, Lycorhinus e Pegomastax. Até recentemente, Massospondylus foi considerado como o único sauropodomorph conhecido da Formação Superior Elliot. No entanto, descobertas mais recentes revelaram uma fauna diversa sauropodomorph contemporâneos, com seis espécies adicionais, incluindo os prossaurópodes Ignavusaurus, Arcusaurus e dois táxons, bem como dois saurópodes identificadas.
Não está claro qual carnívoros podem ter predados Massospondylus. A maioria dos terópodes que foram descobertas em rochas de idade jurássico adiantado na África Austral, como Megapnosaurus, foram menores do que prossaurópodes de médio porte como Massospondylus. Esses predadores menores têm sido postulados como usar ataques rápidos cortando a desgastar prossaurópodes, que poderia ter-se defendido com as suas mãos e os pés grandes garras. A 6-metre- (20 FT) terópode carnívoro longo Dracovenator viveu durante o mesmo período (Hettangian para estágios Sinemurian) como Massospondylus e também foi encontrado na Formação Elliot da África do Sul.
Paleobiology
Tal como acontece com todos os dinossauros, em muito da biologia de Massospondylus, incluindo o seu comportamento, coloração, e fisiologia, permanece desconhecida. No entanto, estudos recentes têm permitido a especulação informada sobre assuntos como os padrões de crescimento, dieta, postura, reprodução e respiração.
Um estudo de 2007 sugeriu que Massospondylus pode ter usado seus braços curtos na defesa contra predadores ("swats defensivas"), em combate intra-espécies, ou na alimentação, embora seus braços eram muito curtos para alcançar a sua boca. Os cientistas especulam que Massospondylus poderia ter usado sua grande pollex (polegar) garra em combate, para retirar material de plantas de árvores, cavando, ou para grooming.
Crescimento

Um estudo de 2005 indicou que Massospondylus 'irmã taxon, Plateosaurus, exibiram padrões de crescimento afetadas por fatores ambientais. O estudo indicou que quando a comida era abundante, ou quando o clima foi favorável, Plateosaurus apresentaram crescimento acelerado. Este padrão de crescimento é chamado de "plasticidade do desenvolvimento". É invisível em outros dinossauros, incluindo Massospondylus, apesar da estreita relação entre Plateosaurus e Massospondylus. O estudo indicou que Massospondylus cresceu ao longo de uma trajectória de crescimento específico com pouca variação na taxa de crescimento e o tamanho final de um indivíduo. Outro estudo de determinação da idade indicaram que Massospondylus cresceu a uma taxa máxima de 34,6 kg (76,3 £) por ano e ainda estava crescendo em torno de 15 anos de idade.
Dieta
Prossaurópodes como Massospondylus pode ter sido herbívoros ou onívora. Tão recentemente quanto 1980, os paleontólogos debatida a possibilidade de carnivory em prossaurópodes. No entanto, a hipótese de prossaurópodes carnívoras foi desacreditada, e todos os estudos recentes favorecem um estilo de vida herbívoros ou onívoros para estes animais. Galton e Upchurch (2004) verificaram que as características cranianos (tais como mandíbula articulação) da maioria dos prossaurópodes estão mais próximas das dos répteis herbívoros do que aqueles de uns carnívoros, e a forma do dente coroa é semelhante aos dos herbívoros ou omnívoros moderna iguanas. A largura máxima da coroa foi maior do que a da raiz, resultando em uma aresta de corte semelhantes aos de répteis herbívoros ou omnívoros existentes. Barrett (2000) propôs que prossaurópodes suplementado suas dietas herbívoros com pequenas presas ou carniça. Gastrólitos (pedras de moela) foram encontrados em associação com três fósseis Massospondylus da Mata-Sandstone no Zimbabwe , e com um animal -como Massospondylus do Triássico Superior de Virginia. Até recentemente, os cientistas acreditam que estas pedras funcionava como um moinho gástrico para auxiliar a ingestão de material vegetal, para compensar a sua incapacidade para mastigar, como é o caso em muitas aves modernas. No entanto, Asas e Sander (2007) mostrou que a natureza polida ea abundância dessas pedras impedido um uso como um moinho gástrico eficaz na maioria dos não- terópodes dinossauros, incluindo Massospondylus.
Marcha e amplitude de movimento

Embora longo assumiu ter sido quadrúpede, um estudo anatômico do forelimbs 2007 questionou isso, argumentando que a sua gama limitada de movimento impedia a marcha quadrúpede habitual eficaz. Nem os membros anteriores poderia oscilar a frente e para trás de um modo semelhante ao dos membros posteriores, nem pode a mão ser rodado com as superfícies palmar virados para baixo. Esta incapacidade de pronate a mão também é suportado pelo in-situ achados de articulados (ainda conectados) braços que sempre mostram as mãos não giradas com palmar enfrenta frente para o outro. O estudo também descartou a possibilidade de "nodopedalia" e outras formas de locomoção que faria uma locomoção eficaz possível, sem a necessidade de pronação a mão. Apesar de sua massa sugere uma natureza quadrúpede, Massospondylus teria sido restrita a suas pernas traseiras para locomoção.
Desde a descoberta de rudimentar e não funcional clavículas em ceratopsians supunha-se que esses ossos do ombro foram reduzidos em todos os dinossauros que não têm verdadeiro furculae. Robert Bakker (1987) sugeriram que este teria permitido a omoplatas para balançar com os membros anteriores em dinossauros quadrúpedes, aumentando seu comprimento forelimb funcional. Isso teria reduzido a discrepância de comprimento entre previsão e membros posteriores em um Massospondylus quadrúpede. No entanto, uma descoberta recente mostra que possuía Massospondylus bem desenvolvida clavículas que se juntaram em um arranjo de fúrcula-like, agindo como um fecho entre as escápulas direita e esquerda e que proíbe qualquer rotação desses ossos. Esta descoberta indica que a redução da clavícula se limita à linha evolutiva que conduz aos ceratópsios. Ele também indica que o fúrcula de aves é derivado de clavícula.
Michael Cooper (1981) observou que o zigapófises das vértebras do pescoço estavam inclinados, proibindo o movimento horizontal significativa do pescoço, de modo que "consequentemente, qualquer movimento significativo nesse sentido devem ter sido conseguida por uma alteração da posição do corpo inteiro". Este foi contrariada em um estudo recente, notando que apenas os cervicais basalmost mostrar zigapófises inclinadas, permitindo o movimento horizontal suficiente do pescoço como um todo.
Reprodução

Em 1976, uma embreagem de sete de 190 milhões de anos de idade, ovos Massospondylus foram encontrados em Golden Gate Highlands National Park na África do Sul por James Kitching, que os identificados como mais provável que pertence a Massospondylus. Foi quase 30 anos antes da extração foi iniciado nos fósseis do 15-centimetre- (6 in-) embriões longos. Eles continuam a ser os mais antigos embriões de dinossauro já encontrado. No início de 2012, pelo menos 10 desovas de pelo menos quatro horizontes fossilíferos havia sido encontrado, com até 34 ovos por desova. Isto indica que este local de assentamento pode ter sido usado repetidamente (local de fidelidade), por grupos de animais (nidificação colonial); em ambos os casos, estes representam a mais antiga evidência deste comportamento. Estruturas sedimentares indicam que a área de assentamento estava na vizinhança de um lago. Os ovos foram muito fina (cerca de 0,1 mm), permitindo a troca de gás, mesmo em um de oxigénio e dióxido de carbono de baixa ambiente rico, o que indica que os ovos foram pelo menos parcialmente enterrada no substrato. Não há indícios de que Massospondylus ninhos construídos; No entanto, a disposição dos ovos em linhas apertadas indica que os ovos foram empurrado nesta posição pelos adultos.
Os embriões provavelmente representou quase-filhotes. Embora as características do esqueleto foram semelhantes aos dos adultos, as proporções corporais foram muito diferentes. A cabeça era grande com um focinho curto e muito grandes órbitas, cujo diâmetro atinge 39% de todo o comprimento do crânio. O pescoço era curto, contrastando com o pescoço muito longo nos adultos. Cinturão ossos e caudais eram relativamente pequenas. Os membros dianteiros eram iguais para os membros posteriores, indicando que Massospondylus recém-nascidos eram quadrúpedes, ao contrário dos adultos bípedes. A descoberta de pegadas de incubação com impressões manus confirmaram a sua quadrupedality. Essas impressões mostram que a mão não foi pronado, com a palma enfrenta frente para o outro e o polegar virado para frente. O manus unpronated ea grande cabeça indicam que a locomoção eficaz não foi possível para recém-nascidos Massospondylus. Notavelmente, os quase-hatchings não tinha dentes, sugerindo que eles não tinham como alimentar-se. Com base na falta de dentes e a locomoção ineficaz, os cientistas especulam que os cuidados pós-natal pode ter sido necessário. Este é ainda apoiada pela evidência de que os hatchings manteve-se nos ninhos até que dobrou de tamanho.

Recém-nascidos menores são conhecidos a partir de uma segunda prosauropod, Mussauro; esses restos se assemelham às do Massospondylus embrionárias, sugerindo que quadrupedality estava presente em recém-nascidos mussauro e presumivelmente outros prossaurópodes também. O quadrupedality das eclosões sugere que a postura quadrúpede de tarde saurópodes podem ter evoluído a partir de retenção de características juvenis em animais adultos, um fenômeno evolutivo conhecido como pedomorphosis. Esta descoberta, portanto, "lança alguma luz nos caminhos evolutivos através do qual as adaptações peculiares de dinossauros gigantes foram atingidos", afirmou o paleontólogo francês Eric Buffetaut.
Sistema respiratório
Muitos saurischian dinossauros possuíam vértebras e costelas que continham cavidades ocas (pneumático forames), que reduziu o peso dos ossos e pode ter servido como um sistema de base »ventilação de escoamento" parecido com o das aves modernas. Num tal sistema, as vértebras do pescoço e nervuras são escavado pelo saco de ar do colo do útero; as vértebras parte superior das costas, pelo pulmão; e parte inferior das costas e sacral (hip) vértebras, pelo saco de ar abdominal. Estes órgãos, constituem um método complexo e muito eficiente de respiração. Prossaurópodes são o único grande grupo de saurischians sem um sistema extensivo de forame pneumático. Embora possíveis entalhes pneumáticos têm sido encontrados em Plateosaurus e thecodontosaurus, as reentrâncias foram muito pequenas. Um estudo realizado em 2007 concluiu que prossaurópodes como Massospondylus provavelmente teve sacos de ar abdominais e cervicais, com base nas provas para eles na irmã táxons (terópodes e saurópodes). O estudo concluiu que era impossível determinar se prossaurópodes teve um flow-through de pulmão de pássaro, mas que os sacos de ar foram quase de certeza presente.