
Réptil
Informações de fundo
Esta seleção Wikipedia está offline disponível a partir de Crianças SOS, para distribuição no mundo em desenvolvimento. Uma boa maneira de ajudar outras crianças é por patrocinar uma criança
| Répteis Gama Temporal: Carbonífero - Recentes | |
|---|---|
 | |
| A Tuatara, Sphenodon punctatus | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Subfilo: | Vertebrata |
| Classe: | Sauropsida * Goodrich, 1916 |
| Subclasses | |
| |
| Sinônimos | |
| |
Os répteis são ar-respirando, de sangue frio vertebrados que têm pele coberta de Escalas ao contrário de cabelo ou penas. Eles são tetrápodes (tendo ou tendo descido de vertebrados com quatro membros) e amniotes, cujas embriões são rodeados por um amniótico membrana. Répteis modernos habitam cada continente com exceção da Antártica , e são representadas por quatro de estar ordens:
- Crocodilia ( crocodilos , gavials, jacarés e jacarés ): 23 espécies
- Sphenodontia ( tuatara da Nova Zelândia ): 2 espécies
- Squamata ( lagartos , cobras e anfisbenídeos ("verme-lagartos")): cerca de 7900 espécies
- Testudines ( tartarugas e cágados ): aproximadamente 300 espécies
A maioria das espécies de répteis são ovíparo (postura de ovos), embora algumas espécies de squamates são capazes de dar à luz ao vivo. Isto é conseguido, quer através ovoviviparity (retenção de ovos), ou viviparity (descendentes nascidos sem uso de calcificadas ovos). Muitas das espécies vivíparos alimentar a sua fetos através de várias formas de placenta análogas às de mamíferos com algum cuidado inicial prevê a sua filhotes. Répteis existentes variam em tamanho do recém-descoberto Jaraguá Sphaero, a 1,6 cm (0,6 in), com a Crocodilo de água salgada, em até pelo menos 7 m (23 ft).
Classificação
História da classificação
Do ponto de vista clássico, répteis incluído todas as amniotas exceto aves e mamíferos . Assim répteis foram definidos como o conjunto de animais que inclui crocodilos , jacarés , tuatara, lagartos , cobras, anfisbenídeos e tartarugas , agrupados como a classe Reptilia (repere Latina ", a rastejar"). Esta ainda é a definição usual do termo. No entanto, nos últimos anos, muitos taxonomistas têm começado a insistir que taxa deve ser monofiléticas, isto é, os grupos devem incluir todos os descendentes de uma forma particular. Os répteis como definidos acima seria paraphyletic, uma vez que exclui as aves e os mamíferos, embora estes também desenvolvido a partir do réptil originais. Colin Tudge escreve:
| " | Os mamíferos são um clade, e, portanto, os cladistas estão felizes em reconhecer o taxon tradicional Mammalia ; e aves, também, são um clado, universalmente atribuída às formais táxon Aves . Mammalia e Aves são, de fato, subclades dentro da grande clade do Amniota. Mas o reptilia aula tradicional não é um clade. É apenas uma seção do clade Amniota: a seção que é deixado após a Mammalia e Aves foram separadas. Ele não pode ser definido por synamorphies, como é o modo adequado. É em vez definido por uma combinação das características que tem e os recursos que ele não tem: répteis são os amniotas que carecem de pêlos ou penas. Na melhor das hipóteses, os cladistas sugerem, poderíamos dizer que o Reptila tradicional são "não-aviários, amniotes não mamíferos». | " |
| " | Do mesmo modo, a classe tradicional torna-se anfíbios anfíbios *, porque alguns antigo deu origem anfíbio ou outra para todos os amniotes; e crustáceos filo Crustacea * torna-se, porque ele pode ter dado origem aos insetos e miriápodes (centopeias). Se acreditarmos, como alguns (mas não todos) os zoólogos não, que myriapods deu origem a insetos, então eles devem ser chamado Myriapoda * .... por esta convenção Reptilia sem um asterisco é sinônimo de Amniota, e inclui aves e mamíferos, Considerando Reptilia * significa não-aviários, amniotes não mamíferos. | " |
Os termos "Sauropsida" ("Faces do lagarto") e " Theropsida "(" Animal Faces ") foram cunhado em 1916 por ES Goodrich para distinguir entre lagartos, pássaros e seus familiares, por um lado (Sauropsida) e mamíferos e seus parentes extintos (Theropsida), por outro. Goodrich apoiou esta divisão pela natureza dos corações e vasos sanguíneos em cada grupo, e outras características, tais como a estrutura do cérebro anterior. De acordo com a Goodrich, ambas as linhagens evoluíram a partir de um grupo haste anteriormente, os Protosauria ("Primeiro Lagartos") que incluiu alguns Paleozóico anfíbios, bem como primeiros répteis.
Em 1956 DMS Watson observou que os dois primeiros grupos divergiram muito cedo na história dos répteis, e assim ele dividiu Protosauria da Goodrich entre eles. Ele também reinterpretou a Sauropsida e Theropsida excluir aves e mamíferos, respectivamente. Assim, sua Sauropsida incluído Procolophonia, Eosuchia, Millerosauria, Chelonia (tartarugas), Squamata (lagartos e cobras), Rhynchocephalia, Crocodilia, " tecodontes "( parafiléticos basal Archosauria), não- aviário dinossauros , pterossauros , ictiossauros , e sauropyterygians.
Esta classificação suplementado, mas nunca foi tão popular como, a classificação dos répteis (de acordo com Clássico de Romer Vertebrados Paleontologia) em quatro subclasses de acordo com o posicionamento de fenestrae temporal aberturas nos lados do crânio por trás dos olhos. Estas divisões foram:
- Anapsida - não fenestrae
- Synapsida - um baixo fenestra (não mais considerados verdadeiros répteis)
- Euryapsida - uma fenestra elevadas (agora incluídos dentro Diapsida)
- Diapsida - dois fenestrae
Todos os itens acima, mas Synapsida queda sob Sauropsida.
Taxonomia
A classificação para a ordem de nível, depois de Benton, 2004.
- Série Amniota
- Classe Synapsida
- Ordem Pelycosauria *
- Ordem Therapsida
- Classe Mammalia
- Classe Sauropsida
- Subclasse Anapsida
- Ordem Testudines (tartarugas)
- Subclasse Diapsida
- Ordem Araeoscelidia
- Ordem Younginiformes
- Infraclass Ichthyosauria
- Infraclass Lepidosauromorpha
- Superordem Sauropterygia
- Ordem Placodontia
- Ordem Nothosauroidea
- Ordem Plesiosauria
- Superordem Lepidosauria
- Ordem Sphenodontida (tuatara)
- Ordem Squamata (lagartos e cobras)
- Superordem Sauropterygia
- Infraclass Archosauromorpha
- Ordem Prolacertiformes
- Divisão Archosauria
- Subdivisão Crurotarsi
- Superordem Crocodylomorpha
- Ordem Crocodylia
- Superordem Crocodylomorpha
- Subdivisão Avemetatarsalia
- Infradivision Ornithodira
- Ordem Pterosauria
- Superorder Dinosauria
- Ordem Saurischia
- Classe Aves
- Ordem Ornithischia
- Ordem Saurischia
- Infradivision Ornithodira
- Subdivisão Crurotarsi
- Subclasse Anapsida
- Classe Synapsida
Filogenia
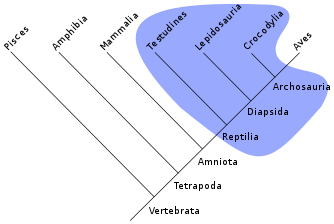
O cladogram aqui apresentado ilustra a "árvore genealógica" de répteis, e segue uma versão simplificada dos relacionamentos encontrados por Laurin e Gauthier (1996), apresentado como parte do Árvore da Vida Project Web.
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Evolução

Hylonomus é o mais antigo conhecido réptil, e foi cerca de 8 a 12 polegadas (20 a 30 cm) de comprimento. Westlothiana tem sido sugerido como o réptil mais antigo, mas é para o momento considerado mais relacionado com anfíbios do que amniotes. Petrolacosaurus e Mesosaurus são outros exemplos. Os primeiros répteis foram encontrados nas florestas pantanosas do Carbonífero , mas foram em grande parte ofuscado pelo maior anfíbios labyrinthodont como Proterogynrius. Foi só depois de a pequena idade do gelo no final do Carbonífero que os répteis cresceu para tamanhos grandes, produzindo espécies como Edaphosaurus e Dimetrodon .
Os primeiros verdadeiros "répteis" (sauropsídeos) são classificados como Anapsids, com um crânio maciço com buracos única para nariz, olhos, medula espinhal, etc. Tartarugas são acreditados por alguns de ser sobrevivendo Anapsids, como eles também compartilham esta estrutura do crânio; mas este ponto tornou-se controversa, ultimamente, com alguns argumentando que tartarugas revertido para este estado primitivo, a fim de melhorar a sua armadura. Ambos os lados têm fortes evidências, eo conflito ainda não foi resolvido.
Logo após os primeiros répteis, dois ramos se separaram, um líder para os Anapsids, que não desenvolveram buracos em seus crânios. O outro grupo, Diapsida, possuía um par de furos nas suas crânios por trás dos olhos, juntamente com um segundo par localizado mais elevado no crânio. O Diapsida dividido novamente em duas linhagens, o Lepidosaurs (que contêm cobras modernas, lagartos e tuataras, bem como, debatably, os répteis extintos mar do Mesozóico) eo arcossauros (hoje representados apenas por crocodilianos e aves com menos de dinossauros , mas também contendo pterossauros e dinossauros não-aviários).
O mais antigo, solid-skulled amniotes também deu origem a uma linha separada, a Synapsida. Sinapsídeos desenvolvido um par de furos nos seus crânios por trás dos olhos (semelhantes às diapsids), que foram utilizados para iluminar ambos o crânio e aumentar o espaço para os músculos do maxilar. Os synapsids eventualmente evoluiu para os mamíferos , e são muitas vezes referida como répteis semelhantes a mamíferos, embora eles não são verdadeiros membros da Sauropsida. (Um termo preferido é "tronco-mamíferos".)
Sistemas
Circulatório

A maioria dos répteis têm um de três câmaras do coração que consiste em dois átrios, uma variável-particionado ventrículo, e dois aorta que ir a do circulação sistémica. O grau de mistura de oxigenado sangue desoxigenado e no coração de três câmaras é variável, dependendo da espécie e estado fisiológico. Sob diferentes condições, o sangue venoso pode ser desviado de volta para o corpo ou sangue oxigenado pode ser desviado de volta para os pulmões. Esta variação no fluxo sanguíneo foi levantada a hipótese de permitir termorregulação mais eficaz e tempos de mergulho mais longos para espécies aquáticas, mas não foi demonstrado ser um vantagem de fitness.
Há algumas exceções interessantes para a fisiologia geral. Por exemplo, crocodilianos têm um coração de quatro câmaras anatomicamente, mas também tem dois aorta sistêmica e, portanto, são capazes apenas de contornar a sua circulação pulmonar. Além disso, algumas cobras e lagartos espécies (por exemplo, monitor lagartos e as pítons) têm corações três câmaras que se tornam corações funcionais quatro câmaras durante a contração. Isto é possível por uma crista muscular que divide o ventrículo durante diástole ventricular e divide-o completamente durante sístole ventricular. Devido a este cume, alguns deles squamates são capazes de produzir os diferenciais de pressão ventriculares que são equivalentes aos observados em corações de mamíferos e aves.
Respiratório
Todos os répteis respirar usando pulmões. Aquáticos tartarugas desenvolveram pele mais permeável, e algumas espécies têm modificado sua cloaca para aumentar a área para a troca gasosa (Orenstein, 2001). Mesmo com essas adaptações, a respiração nunca é completamente realizado sem pulmões. Ventilação pulmonar é realizada de forma diferente em cada grupo réptil principal. Em squamates os pulmões são ventilados quase exclusivamente pela musculatura axial. Este é também o mesmo musculatura que é usado durante a locomoção. Devido a esta restrição, a maioria dos squamates são obrigados a segurar a respiração durante as corridas intensas. Alguns, no entanto, ter encontrado uma maneira de contornar isso. Varanids, e algumas outras espécies de lagartos, empregam bombeamento bucal como um complemento ao seu normal "respiração axial." Isto permite que os animais para encher completamente os pulmões durante a locomoção intensa, e assim permanecem aerobicamente activa durante um longo tempo. Lagartos Tegu são conhecidos por possuir um proto diafragma, o qual separa a cavidade pulmonar da cavidade abdominal. Embora não seja efectivamente capaz de se movimentar, ele permite uma maior inflação pulmonar, tomando o peso das vísceras fora dos pulmões (Klein et al, 2003). Crocodilians realmente ter um diafragma muscular que é análogo ao diafragma de mamífero. A diferença é que os músculos para o diafragma de crocodilo puxar o púbis (parte da pélvis, que é móvel em crocodilians) para trás, o que traz o fígado para baixo, libertando assim espaço para os pulmões de expanda. Este tipo de configuração diafragmática tem sido referido como o "pistão hepática."
Como tartarugas e cágados respirar tem sido objecto de muito estudo. Até à data, apenas algumas espécies foram estudadas cuidadosamente o suficiente para ter uma idéia de como as tartarugas fazê-lo. Os resultados indicam que as tartarugas e jabutis têm encontrado uma variedade de soluções para este problema. O problema é que a maioria dos cascos de tartaruga são rígidas e não permitem o tipo de expansão e contração que outros amniotes usar para ventilar os pulmões. Algumas tartarugas, como o flapshell indiana (Lissemys punctata) tem uma folha de músculo que envolve os pulmões. Quando se contrai, a tartaruga pode exalar. Quando em repouso, a tartaruga pode retrair os membros na cavidade do corpo e forçar o ar para fora dos pulmões. Quando a tartaruga prolonga as suas extremidades, a pressão no interior dos pulmões é reduzida, e a tartaruga pode sugar o ar no. Tartaruga pulmões são fixados à face interior da parte superior do reservatório (carapaça), com a parte inferior dos pulmões associadas (através conjuntivo tecido) para o resto das vísceras. Usando uma série de músculos especiais (aproximadamente equivalente a um diafragma), as tartarugas são capazes de empurrar a sua vísceras cima e para baixo, resultando em respiração eficaz, uma vez que muitos desses músculos têm pontos de fixação em conjunto com os seus membros anteriores (na verdade, muitos dos músculos expandir para os bolsos dos membros durante contração). A respiração durante a locomoção foi estudado em três espécies, e mostram padrões diferentes. Feminino tartarugas marinhas verde adultos não respirar como eles muleta ao longo de suas praias de desova. Eles segurar a respiração durante a locomoção terrestre e respirar em ataques como eles descansam. North American caixa tartarugas respirar continuamente durante a locomoção, eo ciclo de ventilação não está coordenada com os movimentos dos membros (Landberg et al., 2003). Eles provavelmente estão usando seus músculos abdominais para respirar durante a locomoção. A última espécie ter sido estudado é vermelho-eared sliders, que também respiram durante a locomoção, mas eles tiveram respirações menores durante a locomoção do que durante pequenas pausas entre as lutas de locomoção, indicando que pode haver interferência mecânica entre os movimentos dos membros e do aparelho respiratório. Tartarugas de caixa também foram observados para respirar enquanto completamente selado dentro de suas conchas (ibid).
A maioria dos répteis não têm um palato secundário, o que significa que eles devem prender a respiração ao engolir. Crocodilianos têm evoluído um palato secundário ósseo que lhes permite continuar a respiração enquanto permanece submerso (e proteger seus cérebros de ser chutado em lutando presa). Skinks (família Scincidae) também desenvolveram um palato secundário ósseo, em graus variados. Snakes teve uma abordagem diferente e ampliou sua traqueia em vez disso. Sua extensão traqueal sobressai como uma palha carnuda, e permite que estes animais de engolir grandes presas, sem sofrer de asfixia.
Excretório
Excreção é realizada principalmente por dois pequenos rins . Em diapsids ácido úrico é o principal nitrogenados produto de resíduos; tartarugas, como mamíferos , excretam principalmente ureia . Ao contrário dos rins de mamíferos e aves, répteis rins são incapazes de produzir líquido de urina mais concentrada do que o seu fluido corporal. Isto é porque lhes falta uma estrutura especializada presente nos nefrónios de mamíferos e aves, um chamado Alça de Henle. Devido a isso, muitos répteis usar o cólon para auxiliar na reabsorção de água. Alguns também são capazes de absorver a água armazenada no bexiga. Excesso de sais também são excretados por nasais e lingual de sal-glândulas em alguns répteis.
Nervoso
O sistema nervoso reptiliano contém a mesma parte básica do anfíbio cérebro, mas o réptil cérebro e cerebelo são ligeiramente maiores. A maioria dos órgãos dos sentidos típicos são bem desenvolvidos com algumas excepções mais notavelmente o cobras falta de orelhas externas (orelhas média e interna estão presentes). Todos os reptilianos têm avançado percepção profundidade visual em comparação com outros animais. Há doze pares de nervos cranianos.
Reprodutivo
A maioria dos répteis se reproduzem sexualmente, embora alguns são capazes de reprodução assexuada. Toda a atividade reprodutiva ocorre com a cloaca, o único saída / entrada na base da cauda, onde os resíduos são eliminados. Tuataras falta órgãos copulatórios, de modo que o macho e fêmea basta pressionar suas cloacas juntas como o macho excreta esperma. A maioria dos répteis, no entanto, têm órgãos copulatórios, os quais são normalmente retraídos ou invertidos e armazenados no interior do corpo. Em tartarugas e crocodilos, o macho tem uma única média pénis, enquanto squamates incluindo cobras e lagartos possuem um par de hemipenes.
A maioria dos répteis botam ovos amniótico cobertas com couro ou calcários conchas. Um amnion, e chorion alantóide estão presentes durante vida embrionária. Não há estágios larvais de desenvolvimento. Viviparity e ovoviviparity única evoluíram em squamates, e uma fração substancial das espécies utilizam este modo de reprduction, incluindo todas as boas ea maioria das víboras. O grau de viviparity varia: algumas espécies simplesmente manter os ovos até pouco antes de chocar, outros fornecem alimento materno para complementar a gema, enquanto outros ainda não têm qualquer gema e fornecer todos os nutrientes através de uma placenta.
A reprodução assexuada foi identificada em squamates em seis famílias de lagartos e uma cobra. Em algumas espécies de squamates, uma população de fêmeas são capazes de produzir um clone diplóide unissexual da mãe. Esta reprodução assexuada chamado partenogénese ocorre em várias espécies de gecko, e é particularmente comum no teiids (especialmente Aspidocelis) e (lacertídeos Lacerta). Em cativeiro Os dragões de Komodo (Varanidae) reproduziram por partenogênese.
Espécies partenogenéticas também são suspeitos de ocorrer entre camaleões, agamids, xantusiids, e typhlopids.